


 我要投稿
我要投稿
介绍
慢性疼痛状况是全世界最常见的致残原因之一。除了疼痛和残疾以外,慢性疼痛还与认知和情绪障碍有关,这进一步降低了生活质量。由于疗效和不良反应,当前的药物治疗常常不能满足患者的需求。另外,合并症可能导致患者寻求多种治疗,增加了他们的经济负担。因此,必须采取针对慢性疼痛的这些多维方面的整体方法,同时控制副作用,以增强治疗效果.迄今为止,人类研究表明,包括海马,前扣带回皮层,内侧前额叶皮层和背侧缝核在内的棘上神经结构与慢性疼痛有关。特别是在慢性疼痛患者中发现海马体积减少和海马结构改变,这意味着与疼痛相关的认知功能障碍。许多研究还显示,在慢性疼痛的动物模型中,海马功能受损,包括异常的细胞因子表达,短期和工作记忆缺陷以及长期增强(LTP)受损。
神经损伤还影响突触可塑性,并通过海马兴奋性和抑制性传递的表达或功能改变来诱导多种神经递质和细胞内信号转导的大量变化。谷氨酸是中枢神经系统的主要兴奋性神经递质,最近的研究已经研究了海马谷氨酸能系统在疼痛的病理生理中的潜在作用。众所周知,谷氨酸通过NMDA受体(NMDAR)和α-氨基-3-羟基5-甲基-4-异恶唑丙酸受体(AMPAR)调节海马兴奋性突触传递。NMDAR亚基NR2B通过调节LTP诱导参与各种生理过程,包括学习,记忆和突触可塑性。此外,钙/钙调蛋白依赖性蛋白激酶II(CaMKII)在LTP诱导中起关键作用。在神经性疼痛模型的海马区发现了谷氨酸受体和CaMKII的水平降低。
临床和动物研究表明,针灸显著改善慢性疼痛。此外,针灸可以减轻与疼痛相关的合并症,如慢性疼痛患者的抑郁和失眠。多项动物研究表明,针刺可改善眼镜蛇毒诱发的慢性神经性疼痛模型中的伤害性和认知相关行为。但是,针灸如何同时挽救合并症和疼痛仍然难以捉摸。
在本研究中,我们首先检查了针刺是否可以改善部分坐骨神经结扎(PSNL)诱发的神经性疼痛模型中的认知和疼痛行为。接下来,使用机器学习中的随机决策森林分类器来查看是否可以基于疼痛和认知行为来预测针灸的治疗效果。然后,我们检查了海马中谷氨酸能受体和突触蛋白以及LTP的变化,以阐明针灸治疗作用的分子机制。
INTRODUCTION
材料和方法
Materials and Methods
动物
将七周大的雄性C57BL / 6小鼠(体重22至25 g;Samaco,韩国首尔)和四周大的雄性C57BL / 6小鼠(用于膜片钳记录;Samtaco)分别放置在24岁在12/12小时的明/暗周期(明:08:00至20:00,暗:20:00至08:00)下在至少2天的时间内保持±2°C,然后自由进行食物和水的实验。
PSNL诱导神经性疼痛
根据Malmberg和Basbaum(1998)的方法建立了左后爪PSNL模型,并进行了一些修改。将小鼠用隆鼻(100μl,腹膜内(ip);Bayer,韩国首尔)和2%zoletil(100μl,ip;Virbac SA,Carros,法国)麻醉。将双侧后腿剃光,并用剪刀暴露坐骨神经。然后,用8-0丝(AILEE,釜山,韩国)将神经背侧的1/3至1/2轻扎,并闭合开放性伤口。在非神经损伤组(假手术;n = 10),神经暴露无损伤并闭合。PSNL手术后7天,将小鼠随机分为5组之一:PSNL,针刺1(AP1),针刺2(AP2),控制点(CP)或阿米替林组(每组)(n = 10)。
针灸与控制疗法
针刺治疗分别在GB30和GB34(AP1组),HT7和GV20(AP2组)或两侧两个控制点(CP组)进行,从术后第7天开始连续连续28天每天进行针刺治疗(图1A)。GB30(在大转子的外侧1/3和内侧2/3与with骨裂孔的交界处)和GB34(在腓骨头的前部和远端的凹陷处)是用于手术的穴位组合木材神经性疼痛的临床治疗。HT7(手掌折痕上的尺骨腕屈肌腱的径向)和GV20(头部的顶点)是经常用于治疗认知和情感障碍的穴位组合。穴位的详细位置如图1A所示。。对于针灸治疗,将小鼠轻度固定,然后将无菌的针灸针(长8 mm,直径0.18 mm;Haenglim-seoweon Acuneedle Co.,韩国)向两侧插入3 mm的深度,然后双向旋转以每秒两次旋转的速度持续30 s,用裸露的手指顺时针旋转180°,逆时针旋转180°。为了排除针刺的非特异性作用,对控制点(CP组)进行了相同的针刺刺激。这两个控制点位于非穴位,距肩cap骨内侧边界或臀肌尾巴基部的中间约3 mm(图1A)。
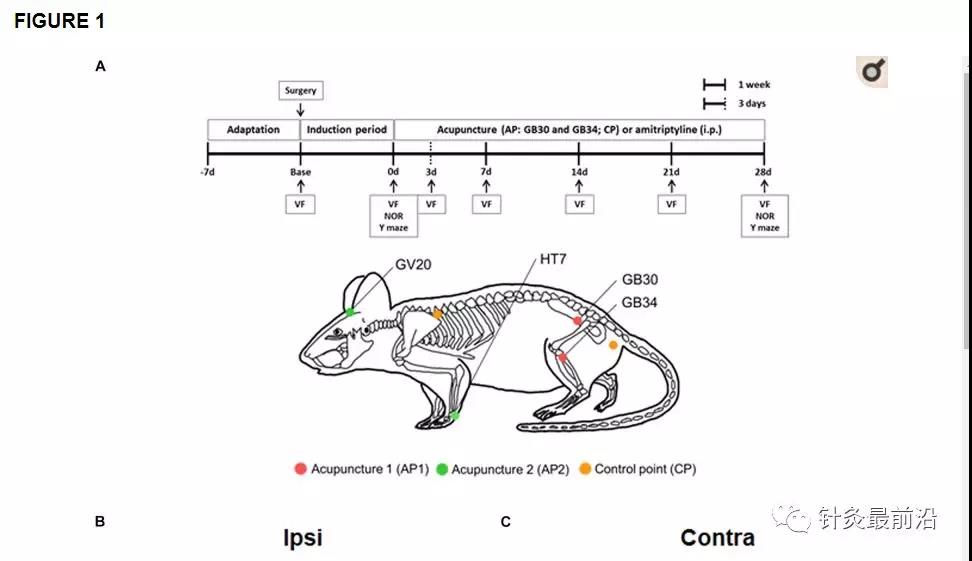
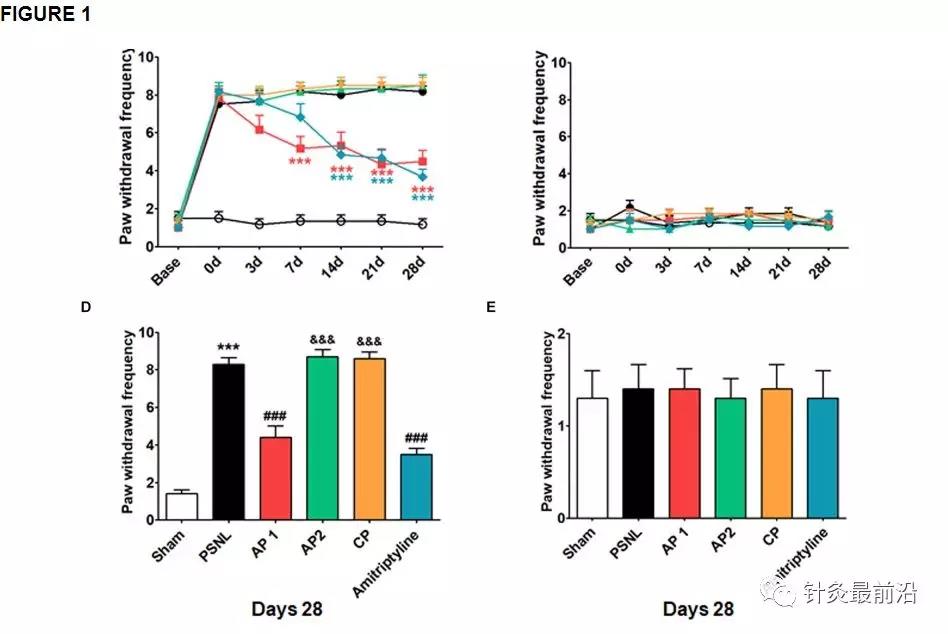
通过von Fey检验评估针灸对机械性异常性疼痛的影响。在PSNL诱发的神经性疼痛模型中施用针灸(AP1,AP2或CP)或阿米替林(10 mg / kg,ip)的实验设计。针灸治疗的位置GB30(Hwando)和GB34(杨凌泉)[AP1(红色圆圈)],HT7(Sinmun)和GV20(Baekhoe)[AP2(绿色圆圈);斑纹绿色圆圈表示背面[back]和控制点[CP(橙色圆圈)] (A)。相对于PSNL组,** p <0.001。给药2小时后,分别测量同侧(B,D)和对侧(C,E)足底表面的针刺(AP1,AP2或CP)或阿米替林的止痛作用(分别为0、3、7、14 , 21天和28天)。ñ= 10 /组。*** p <0.001对Sham组,### p <0.001对所述PSNL基,&&&p <0.001对AP1的基团。数据采用两次重复测量方差分析进行分析,然后进行事后Bonferroni测试。结果表示为平均值±SEM。
阿米替林是一种三环抗抑郁药,被广泛用于治疗各种神经性疼痛。作为阳性对照,从PSNL手术后第7天起,连续28天每天ip给予阿米替林(阿米替林组;100 mg中的10 mg / kg,Sigma-Aldrich,美国密苏里州圣路易斯)。每个治疗日在0.9%NaCl中新鲜制备阿米替林溶液。为了使动物处于相同的压力条件下,Sham,PSNL和阿米替林组动物也被轻度固定,如AP和CP组一样。
伤害行为测试
进行电子冯弗雷测试(IITC,美国加利福尼亚州伍德兰希尔斯)以评估机械性异常性疼痛。这项测试分别在手术和治疗之前进行,并且还在针灸和阿米替林治疗2小时进行。
认知功能测验
通过Y-迷宫中的自发交替行为来测量空间工作记忆。将每只小鼠放在Y迷宫的一只手臂上,并允许他们在5分钟的时间内通过迷宫自由探索。记录输入的武器的顺序和总数。当两只后爪都在手臂上时,手臂进入被认为是完整的。在每次通过之间,用水和乙醇清洁该设备。自发交替的百分比由包含进入所有三个臂的条目的黑社会的数量/最大可能的交替(输入的臂总数-2)×100确定)。
新型对象识别(NOR)测试用于评估短期记忆。仪器装在一个40×40×27 cm的丙烯酸盒中,白色的墙壁和地板。在试验之间清洁盒子和物品,以消除嗅觉提示。动物在空箱中接受了5分钟的训练,以便习惯于仪器和测试室。二十四小时后,在盒子的5分钟“训练阶段”中,每只小鼠都暴露于两个熟悉的物体(圆形块,直径:4厘米)。接下来,将动物放回盒子中,并在“训练阶段”后24小时内将其暴露于一个新颖的物体(矩形块,4×4×4 cm)以及熟悉的物体,再进行5分钟的“测试阶段” 。测量了探索每个对象所花费的时间。反映短期记忆能力的识别指数计算为探索新物体所花费的时间与总探索时间之比。所有认知功能测试均在治疗前(AP1,AP2,CP或阿米替林)进行,并在治疗第28天进行。从治疗后2小时(上午11:00至下午06:00)进行测试。
机器学习中的随机决策森林分类器
根据这三种行为数据(机械异常性疼痛,Y迷宫和NOR),使用随机森林分类器测试我们是否可以预测哪只小鼠属于哪个实验组。随机森林分类器是一种集成方法,将决策树模型与多个随机性结合在一起,并通过这些树的预测值的平均值进行预测。使用Python的scikit学习包1对实验组标记的三种类型的行为数据进行了分析。分类精度是通过应用四重交叉验证获得的。进行一百次迭代以提取平均值。此外,我们通过重复该过程10,000次来随机排列实验组的标签,以生成统计空模型进行比较。为了表达分类的过程和含义,在四个深度级别获得了决策树。最后,使用两对疼痛和认知功能的测量值在散点图上表示鼠标数据。
蛋白质印迹
麻醉后,拔出大脑。将包括海马在内的脑组织样品在200μL裂解缓冲液中匀浆,裂解缓冲液包含20 mM羟乙基哌嗪乙烷磺酸(pH 7.5),1%NP-40、10%甘油,150 mM NaCl,60 mM B-葡糖苷,1 mM苯基甲烷磺酰氟,0.7μg/ mL抑肽酶,磷酸酶和蛋白酶抑制剂混合物片。进行了蛋白质印迹分析以检测海马中谷氨酸能受体和突触蛋白的蛋白表达水平。一抗为兔抗磷酸-NR2B(pNR2B),兔抗-总-NR2B(tNR2B)(按1:1,000稀释;德国默克公司,默克密理博(Merck Millipore),兔抗-磷酸GluR1(pGluR1),兔抗-总-GluR1(tGluR1),兔抗磷酸CaMKII(pCaMKII),兔抗全CaMKII(tCaMKII),兔抗磷酸蛋白激酶C-γ(pPKC-γ),兔抗全蛋白激酶C -γ(pPKC-γ),兔抗磷酸Syn-1(pSyn-1),兔抗全Syn-1(tSyn-1),兔抗磷酸-PSD-95(pPSD-95),兔抗-total-PSD-95(tPSD-95)(按1:1,000稀释;Cell Signaling Technology,美国马萨诸塞州贝弗利)和兔抗-β-肌动蛋白(按1:10,000稀释;Sigma-Aldrich)。然后,将膜与辣根过氧化物酶偶联的山羊抗兔二抗(1:1,000稀释;Pierce,Rockford,IL,美国)一起孵育。使用化学发光试剂盒(Super Signal West Pico;Pierce)将膜可视化,并通过光密度测定法和图像QI软件分析信号强度。我们显示了对比度修改之前的所有波段(补充图4)
免疫荧光
进行了免疫荧光以测量NR2B和GluR1在PFC中神经元细胞中的表达。针对NeuN(小鼠,1:500,MAB377; Chemicon International,Inc.,Temecula,CA,美国),GFAP(小鼠,1:500,14-9892-82; Thermo Fisher Scientific,圣地亚哥,CA)产生的一抗,美国),Iba-1(鼠标,1:500,MABN92;默克密理博),NR2B(兔子,1:500、06-600;默克密理博)和GluR1(兔子,1:1000,#8084;Cell)信号技术)在补充有0.1%BSA的1×PBST中稀释。温育在黑暗中于4℃下进行72小时。接下来,在PBST洗涤后,将切片与Alexa 488偶联的驴抗兔二抗(1:1000; Thermo Fisher Scientific)和Alexa 594偶联的驴抗小鼠二抗(1:1000;Thermo Fisher Scientific)。海马CA1,CA3和DG中NeuN / NR2B和NeuN / GluR1双阳性细胞的数量由研究人员对每组盲人使用正方形网格(300×300μm)进行了三次计数。平均计数定义为NeuN / NR2B和NeuN / GluR1双阳性细胞的数目。此外,将未加抗孵育的组织用作阴性对照(补充图3)。
电生理学
手术后第35天后,进行电生理记录。进行电生理学以测量LTP。异氟烷麻醉后,取出大脑,并用玻璃纤维刀(VT 1200 S;Leica Microsystems,Wetzlar,德国)将海马迅速切开。海马切片(400μm以下)在20mL的人工脑脊液(ACSF孵育;含有119 mM的NaCl的,2.5mM的氯化钾,1mM的的NaH 2 PO 4,26.2毫的NaHCO 3,11 mM的d葡萄糖,2.5mM的氯化钙2,和9 mM MgCl 2;pH = 7.2–7.4;4°C)饱和,含5%CO在32°C的2 in O 2中放置至少1 h。然后,将制备的切片转移至记录室中,含有含氧aCSF中(119毫的NaCl,2.5mM的氯化钾,1mM的的NaH 2 PO 4,26.2毫的NaHCO 3,11 mM的d葡萄糖,2.5mM的氯化钙2,1.3毫摩尔MgCl 2,和0.4 mM抗坏血酸;pH = 7.2–7.4;4°C)在32°C下以2 mL / min的流速流动。玻璃电极(记录电极)填充有记录aCSF,并置于CA1的层半径中。使用微操纵器(ROE-200,Sutter Instruments,Novarto,CA,美国)和控制器(MPC-200,Sutter Instruments)将电极降低至切片表面下方75至150μm。然后,在CA3区放置一个双极电极(刺激电极),以引起田间兴奋性突触后电位(fEPSP)响应。每30秒以0.5 mV / 0.5 ms刺激海马切片至少10分钟,然后以0.5 mV / 100 Hz / 1 s刺激15分钟以进行测试刺激。然后,将由放大器(MultiClamp 700B,Molecular Devices)放大的信号通过A / D转换器(Digidata 1440A,Molecular Devices,Sunnyvale,CA,美国)发送到计算机,并使用Clampex 10.7软件(Molecular Devices)进行分析。
统计分析
所有统计参数均使用GraphPad Prism 5.0软件(GraphPad Software),美国加利福尼亚州圣地亚哥)进行计算。使用未配对的两尾t检验比较假手术组和PSNL组之间在情绪和认知障碍方面的差异。对基于Y迷宫和NOR检验,电生理学,免疫印迹和免疫组化数据的认知障碍行为进行单向方差分析(ANOVA),然后进行Newman-Keuls检验。使用双向重复测量方差分析和Bonferroni post hoc对不同时间点的机械性异常性疼痛进行分析测试成对的多重比较。进行Spearman秩相关系数检验,分析von Frey检验是否与Y-maze和NOR检验相关,海马NR2B和GluR1表达水平是否与von Frey和NOR检验相关。所有数据均表示为平均值±平均值的标准误(SEM)。对于所有分析,认为p <0.05表示具有统计学意义。
结果
针刺可改善PSNL诱发的神经性疼痛的机械性异常性疼痛。
在PSNL诱发的神经性疼痛模型中,使用von Frey检验测量了针灸的抗痛觉和镇痛效果。在PSNL手术前1天进行基线测量。然后,将小鼠随机分为五组。五组的爪子退缩频率没有显示任何差异。在PSNL诱发的神经性疼痛模型中研究了针灸(AP1,AP2或CP)或阿米替林(10 mg / kg,腹膜内)治疗28天的效果(图1)。对于机械性异常性疼痛,采用两次重复测量的方差分析显示组显着效果(F 5,246 = 175.5,p <0.0001)和显着组×时间相互作用(F 30,246 = 8.733,p<0.0001)。Bonferroni 事后测试表明,AP1治疗可在每个时间点逆转已建立的同侧机械性异常性疼痛(治疗后第7天,每个p <0.001 vs. PSNL;图1B)。同样,阿米替林组的爪撤回频率显着低于PSNL组(治疗后第14天,与PSNL 相比,每个p <0.001;图1B)。在第28天,我们还观察到针刺(AP1,AP2或CP)或阿米替林在伤害感受行为测试中的作用。单向方差分析表明,第28天两组之间的缩爪频率存在显着差异(F 5,54 = 68.08,p<0.0001)。Newman–Keuls 事后测试显示,AP1组(p <0.001)和阿米替林组(p <0.001)的爪子退缩频率低于PSNL组(图1D)。对侧后爪的疼痛程度无明显变化(图1C,E)。我们的结果表明,针灸治疗AP1具有镇痛作用。
针刺对PSNL诱发疼痛模型认知功能的影响
为了检查针刺治疗对PSNL诱导的认知障碍的影响,我们使用了Y迷宫和NOR检验(图2)。首先,我们在PSNL手术后8天观察到认知障碍。未配对的两尾t检验显示PSNL小鼠的自发交替显着低于假小鼠(t 18 = 1.120,p = 0.0176;图2A)。此外,未配对的两组t检验表明PSNL手术使PSNL组的识别指数显着降低(t 18 = 12.20,p = 0.0087;图2E)。接着,连续28天继续进行针灸(AP1,AP2或CP)或阿米替林(10 mg / kg,ip)治疗,并在第28天再次测量治疗对认知障碍的影响。在Y迷宫测试中,单向方差分析显示自发轮换组之间存在显着差异(F 5,54 = 8.450,p <0.0001),纽曼- 基尔斯事后检验表明,AP1组的自发轮换高于PSNL组(p <0.05;图2B)。Y迷宫测试的总手臂输入没有明显变化(图2C,D)。在NOR测试中,单向方差分析(F 5,54 = 15.78,p<0.0001),随后进行纽曼(Newman-Keuls)事后检验,结果表明,AP1和阿米替林治疗可恢复PSNL引起的识别指数降低(p <0.001 vs Sham组)(每个p <0.001 vs. PSNL组),而AP2和CP没有影响(图2F)。此外,自发交替和识别指数与爪退缩频率呈负相关(r = -0.6180,p <0.0001;r = -0.6487,p = 0.0026;图2G,H)。因此,我们的结果表明,诱发的神经性疼痛会损害认知功能,而针灸有可能改善神经性疼痛的认知障碍。
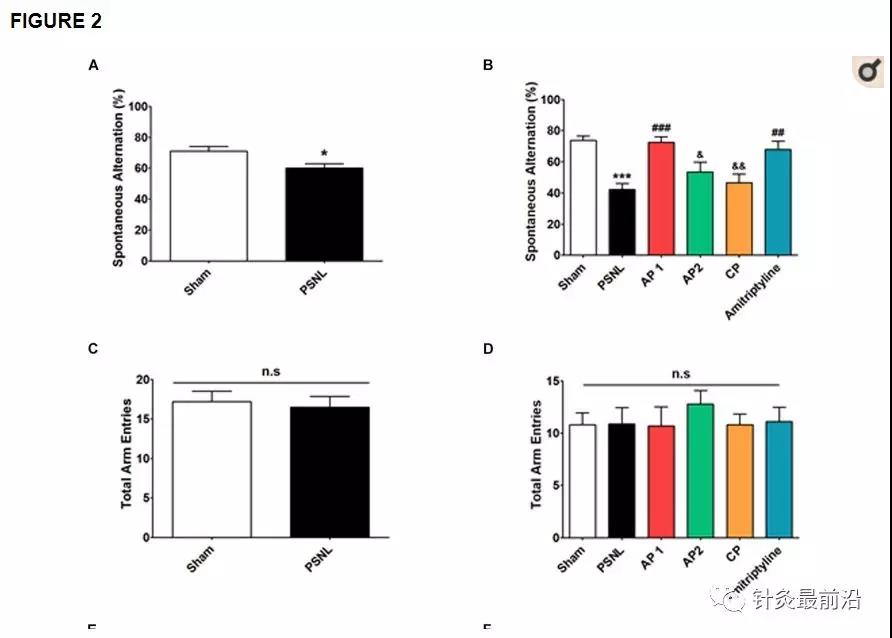
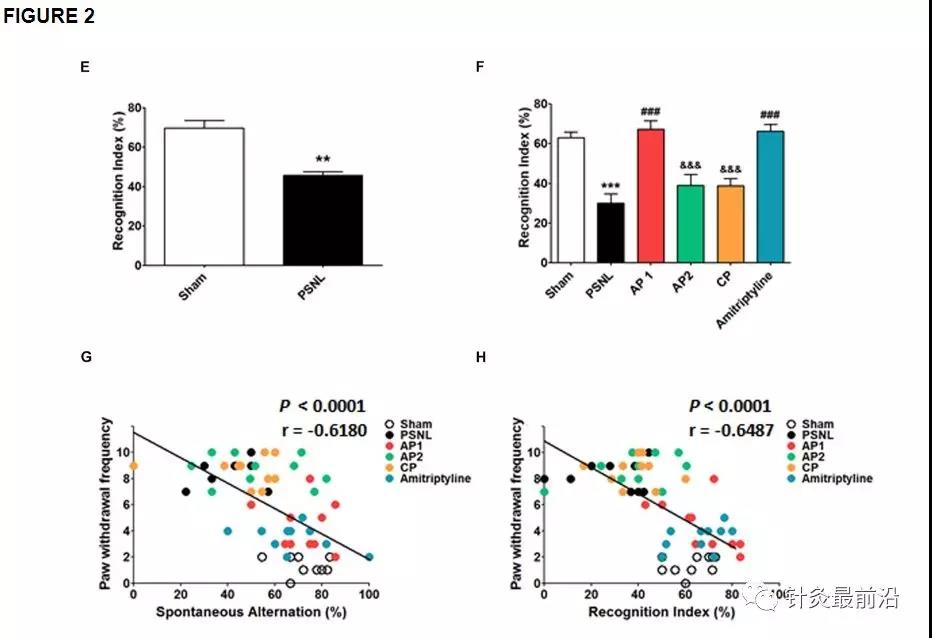
通过Y迷宫和NOR测试,针刺对认知功能的影响。在PSNL诱发的神经性疾病中,使用Y迷宫(A–D)和NOR (E,F)测试研究了针刺(AP1,AP2或CP)或阿米替林(10 mg / kg,ip)对认知障碍的影响疼痛模型。所有组:n =10。** p <0.05,** p <0.01,****** p <0.001 vs. Sham组,#p <0.05,## p<0.01,### p <0.001 vs. PSNL组,&p <0.05,&&p <0.01,和相对于AP1组,p <0.001。对数据进行单向方差分析,然后进行纽曼-基尔斯事后检验(AF)。数据表示为平均值±SEM。Y-迷宫和NOR测验中的认知功能测度与爪退缩频率(G,H)相关。[R -值与Spearman等级相关系数进行分析。AP1,针灸1(GB30和GB34);AP2,针灸2(HT7和GV20);CP,控制点;NOR,新颖的物体识别。
机器学习对针灸对镇痛和认知功能的影响
决策树算法由内部经过培训的随机森林算法组成,以表示行为数据值的分布,分类过程和每个实验组的语义。它在散点图上显示全部40个鼠标数据的决策边界。这示于图3。Sham和AP1组与PSNL和CP组分开。Sham和AP1组显示较低的疼痛反应,而PSNL和CP组显示较高的疼痛反应。疼痛反应数据似乎为区分Sham和AP1组提供了有用的信息,而NOR和Y-迷宫数据则没有。PSNL和CP组之间的疼痛数据没有显着差异,但是Y迷宫和NOR数据显然可以更好地区分它们(图3A,B)。通过应用4倍交叉验证,我们通过疼痛,NOR和Y迷宫这三个阈值以64.4%的准确度预测了每个小鼠在每个实验组中的成员资格。
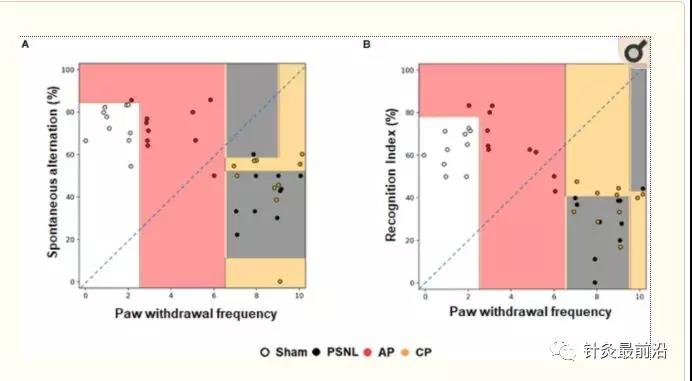
机器学习方法中使用随机森林分类器分析针刺对分类的影响。在机器学习中,通过随机森林算法分析了四组(Sham,PSNL,AP1和CP)的认知功能和伤害感受数据。有代表性的数据表明,针对冯·弗雷测试中的爪子退缩频率,分析了Y迷宫的自发交替(A)和NOR测试(B)的识别指数。所有组:n=10。AP1,针灸1(GB30和GB34);CP,控制点。
针刺对PSNL海马CA3至CA1区LTP的影响
进行LTP记录以研究针灸作用的电生理基础(图4)。针灸治疗(AP1)14-28天后,将fEPSP斜率的时程标准化为基线15分钟,如图4B所示。电刺激后,四组的fEPSP斜率立即增加,并稳定在基线以上的不同水平。最后15分钟的统计平均值如图4C所示。单向方差分析显示,四组之间的fEPSP平均斜率存在显着差异(F 3,120 = 19.17,p <0.001,图4C)。纽曼·基尔斯事后测试显示PSNL和CP组的平均fEPSP斜率比Sham组(p <0.01 vs. PSNL)和AP1组(p <0.001 vs. PSNL组;p <0.001 vs. CP组)小。针灸治疗确实有效逆转了PSNL诱导的LTP损伤。
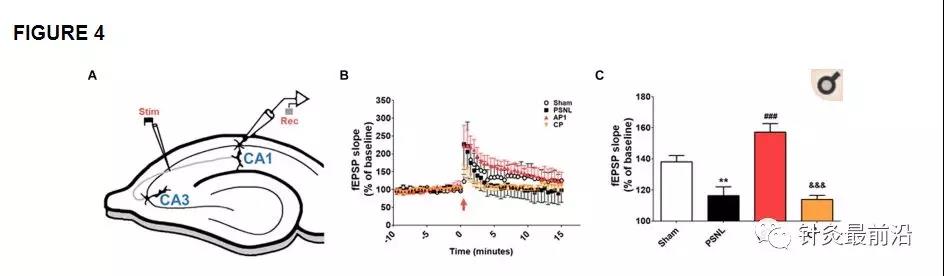
图4
针刺会增加神经性疼痛模型海马CA3至CA1区的fEPSP斜率。在电生理学方法中使用的海马横断面示意图,显示了记录(记录)和刺激(刺激)区域(A)。平均fEPSP数据。在0分钟给予高频刺激,并且信号被跟踪长达15分钟(B)。(C)中显示的fEPSP斜率摘要(从0到15分钟)。所有组中n = 5。** p<0.01 vs. Sham组。与PSNL组相比,### p <0.001,与AP1组相比,&&&p <0.001。对数据进行单向方差分析,然后进行Newman–Keuls分析事后测试。结果表示为平均值±SEM。AP1,针灸1(GB30和GB34);AP2,针灸2(HT7和GV20);CP,控制点。
针刺对PSNL小鼠海马谷氨酸受体表达水平的影响
许多研究表明,NR2B和GluR1在Schaffer侧支CA1突触中通过诱导和维持LTP在突触可塑性中起重要作用。LTP在学习和记忆功能中很重要(Lisman等,2002 ; Fonseca,2012 ; Bliss和Collingridge,2013 ; Wang H.等,2015 ; Shang等,2017)。为了研究谷氨酸受体在神经性疼痛的作用,我们研究海马NR2B和GluR1的采用双免疫染色(图5,6)。6)。首先,对NR2B和GluR1阳性细胞进行双重染色,以检测三种细胞类型标记(GFAP,Iba-1或NeuN)。结果表明它们在神经元中表达,但在星形胶质细胞或小胶质细胞中不表达(补充图1)。接下来,针对针刺组(AP1,AP2或CP)或阿米替林组(10 mg / kg,腹膜内),对海马NR2B和GluR1进行双重免疫染色和蛋白质印迹。对三组(假手术,PSNL和AP1)海马冠状切片进行NR2B和NeuN抗体双重免疫染色。单向方差分析显示NR2B表达组之间存在显着差异(CA1:F 2,7 = 18.26,p = 0.0017; CA2:F 2,7 = 0.2718,p= 0.7697; CA3:F 2.7 = 6.523,p = 0.0252;DG:F 2,7 = 11.23,p = 0.0065),Newman–Keuls 事后检验表明,PSNL组的NR2B表达水平低于Sham组(CA1:p <0.01,CA3:p < 0.05,DG:p<0.01)。AP1给药逆转了PSNL手术导致的海马NR2B表达减弱(CA1:p <0.01,CA3:p <0.05,DG:p <0.01 vs. PSNL; 图5A,B)。在蛋白质印迹分析中,单向方差分析显示两组之间海马中NR2B的蛋白表达存在显着差异(F 5,24 = 6.521,p = 0.0006)。Newman–Keuls 事后测试表明,AP1组的海马NR2B表达水平显着高于PSNL组(p <0.01;图5D)。在海马中,NR2B的表达水平增加与NOR试验的识别指数呈正相关(r = 0.5837和p = 0.0007;图5E),但在von Frey测试中,海马NR2B水平与爪退缩频率之间呈负相关。(r= -0.6403并且p<0.0001;图5F)。
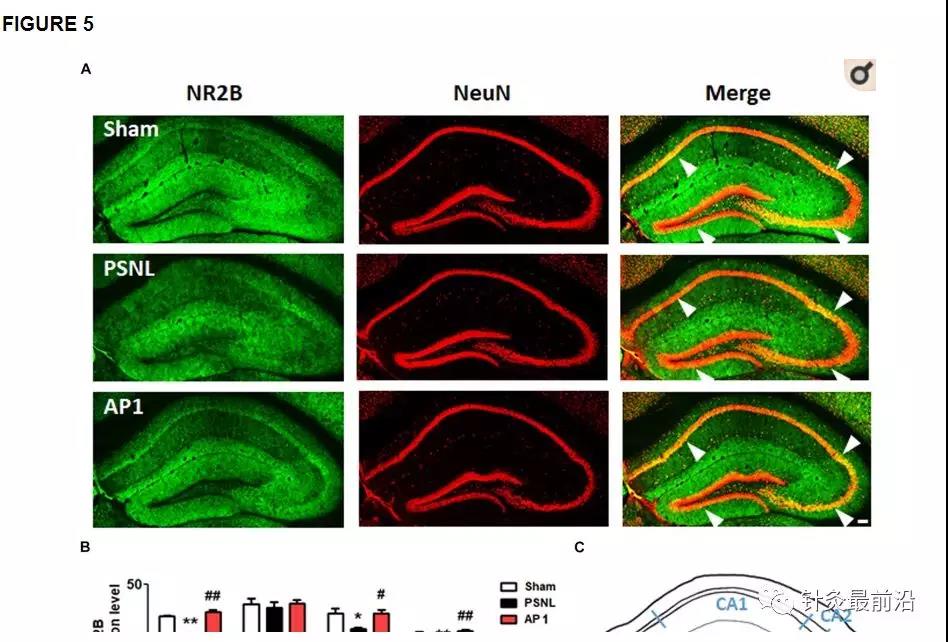
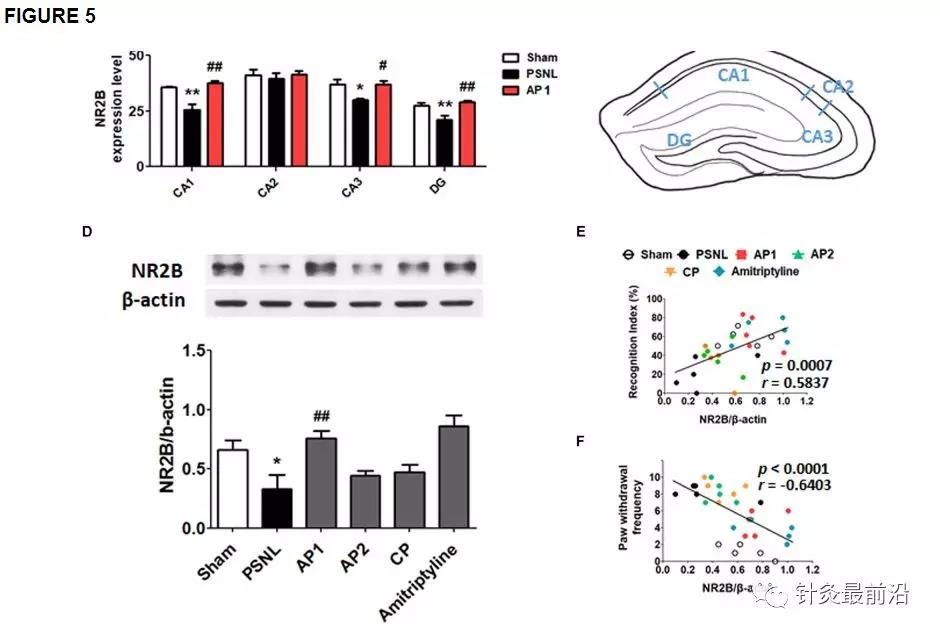
图5
针刺对海马NR2B受体表达水平的影响。这些结果显示,连续28天(AF)施用针灸(AP1,AP2或CP)或阿米替林(10 mg / kg,ip)后,海马NR2B表达水平(CA1,CA2,CA3和DG)的变化)。在PSNL诱导的神经性疼痛模型(A,B)中,AP1给药后海马的组织学检查显示NR2B(绿色)和NeuN(红色)的表达,而在小鼠大脑中的海马区域的代表图(C)。n = 3–4 /组。比例尺:100μm。**p <0.05,** p <0.01,与每个地区的Sham组相比。#与每个区域的PSNL组相比,p <0.05,## p <0.01。在海马中测量NR2B蛋白水平(D)。n = 5 /组。与假手术组相比,∗ p <0.01。与PSNL组相比,#p<0.05。结果通过单向方差分析进行分析,然后进行纽曼-基尔斯事后检验。数据表示为平均值±SEM。NOR测试中的认知功能值和von Frey测试中的伤害感受值与海马NR2B蛋白水平(E,F)相关。在[R值用Spearman等级相关系数进行分析。AP1,针灸1(GB30和GB34);AP2,针灸2(HT7和GV20);CP,控制点。
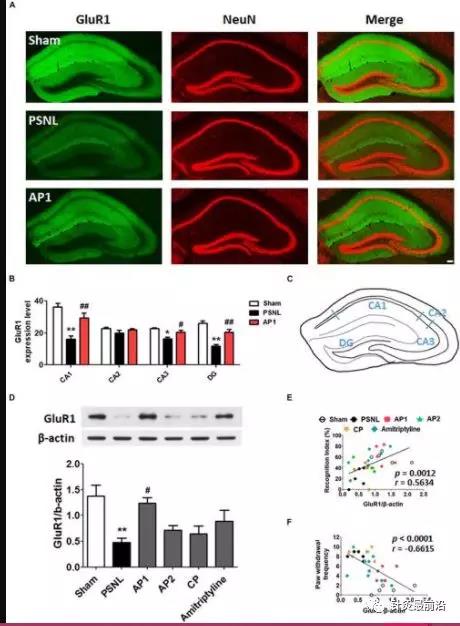
图6
针刺对海马GluR1受体表达水平的影响。这些结果表明,连续28天(AF)施用针灸(AP1,AP2或CP)或阿米替林(10 mg / kg,ip)后海马GluR1表达水平(CA1,CA2,CA3和DG)的变化)。将三组(Sham,PSNL和AP1)的自由浮动冠状海马切片用GluR1(绿色)和NeuN(红色)抗体进行免疫荧光标记,以标记神经元中的GluR1阳性NeuN (A)。代表性图显示了GluR1在海马中的表达水平(A,B),代表性图显示了小鼠脑中的海马区(C)。n = 3–4 /组。比例尺:100μm。** p <0.05,** p<0.01,与每个地区的Sham组相比。#p <0.05,## p <0.01相比于每个区域的PSNL基。检查海马GluR1蛋白的表达水平(D)。n = 5 /组。与假手术组相比,∗ p <0.01。与PSNL组相比,#p <0.05。所有数据均用单因素方差分析后进行纽曼-科伊尔斯分析事后测试。所有数据均表示为平均值±SEM。NOR测试中的认知功能值和von Frey测试中的伤害感受值与海马(E,F)中的GluR1蛋白水平相关。[R -值与Spearman等级相关系数进行分析。AP1,针灸1(GB30和GB34);AP2,针灸2(HT7和GV20);CP,控制点。
接下来,对三组(假手术,PSNL和AP1)的海马冠状切片进行GluR1和NeuN抗体治疗。单向方差分析显示两组之间GluR1的表达存在显着差异(CA1:F 2,7 = 13.27,p = 0.0042; CA2:F 2,7 = 1.900,p = 0.2192; CA3:F 2,7= 6.630,p= 0.0242;DG:F 2.7= 17.98,p= 0.0017)。Newman–Keuls 事后检验表明,PSNL组中的GluR1表达水平低于Sham组(CA1:p <0.01,CA3:p <0.05,DG:p <0.01 vs. PSNL)。类似于NR2B,AP1给药抑制了坐骨神经手术引起的海马GluR1表达水平的降低(相对于PSNL,CA1:p<0.01,CA3:p<0.05,DG:p<0.01;图6A,B)。在蛋白质印迹分析中,单向方差分析显示两组之间GluR1的蛋白表达存在显着差异(F 5,24 = 5.216,p = 0.0022)。Newman–Keuls 事后测试表明,与PSNL组相比,AP1组海马GluR1表达水平显着增加(p<0.01;图6D)。
在NOR试验中,海马中GluR1表达水平与识别指数呈正相关(r = 0.5634和p = 0.0012;图6E),在von Frey试验中与爪退缩频率呈负相关(r = -0.6615和p <0.0001;图6F)。这些结果表明,针刺给药可以通过恢复神经性疼痛中海马中的谷氨酸受体来增强认知功能。
针刺对PSNL小鼠海马CaMKII表达水平的影响
CaMKII和PKC-γ在学习和记忆功能中起关键作用。它们的功能通过突触后膜中的NR2B受体介导的Ca 2+内流来调节。已经在神经性疼痛模型的海马中发现了CaMKII表达水平的降低(Xu等,2012)。因此,我们检查了针刺(AP1,AP2或CP)或阿米替林(10 mg / kg,ip)处理对PSNL小鼠海马中CaMKII表达水平的影响。单向方差分析显示两组之间pCaMKII和tCaMKII的蛋白质表达有显着差异(每个F 5,24 = 10.66; p <0.0001; F 5,24 = 5.156; p= 0.0024),Newman–Keuls 事后检验表明,与假手术组相比,PSNL组的pCaMKII和tCaMKII表达水平降低了(每个p <0.001,p <0.01;图7A,B)。值得注意的是,AP1处理显着恢复了pCaMKII和tCaMKII表达水平(相对于PSNL ,每个p<0.01,p<0.05;图7A,B)。然而,在AP2,CP和阿米替林组中,pCaMKII和tCaMKII蛋白的表达水平没有改善(图7A,B)。通过针刺治疗,海马中的PKC-γ表达水平未改变(补充图2)。
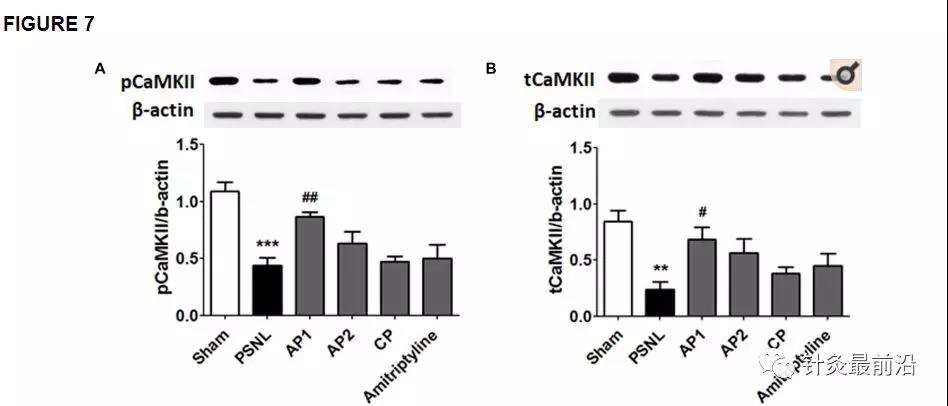
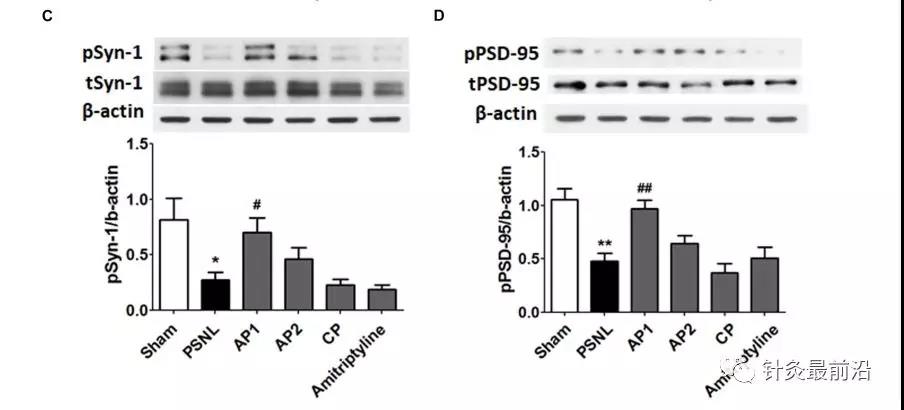
图7
针刺对CaMKII和突触蛋白表达水平的影响。这些结果表明,在针刺(AP1,AP2或CP)或阿米替林给药后,海马CaMKII(A,B;n = 5 /组)和突触蛋白(C,D;n = 4 / group)的蛋白质水平发生了变化。(10 mg / kg,ip)连续28天。** p <0.01,** p * 0.001与Sham组相比。与PSNL组相比,#p<0.05,## p <0.01。数据表示为平均值±SEM。使用相同的β-肌动蛋白值(A,C ; B,D)。AP1,针灸1(GB30和GB34);AP2,针灸2(HT7和GV20);CP,控制点。
针刺对PSNL小鼠海马突触蛋白表达水平的影响
最后,我们研究了针灸对突触蛋白如Syn-1和PSD-95的影响。单向方差分析显示两组之间pSyn-1(F 5,18 = 5.600; p = 0.0028)和pPSD-95(F 5,18 = 10.11; p <0.0001)的蛋白质表达有显着差异,纽曼–Keuls 事后测试表明,与假手术小鼠相比,坐骨神经PSNL引起pSyn-1(p <0.05)和pPSD-95(p<0.01)明显降低。针灸可以显着改善pSyn-1和pPSD-95蛋白水平(每个p <0.05,p相对于PSNL <0.01)。但是,AP2和CP组的pSyn-1和pPSD-95蛋白水平没有明显改变(图7C,D)。我们的数据显示,针刺治疗可恢复神经性疼痛后海马中受损的突触蛋白。
讨论区
慢性疼痛的特征是具有多维方面,涉及伤害感受和情感或认知游行。已知针灸具有镇痛作用并改善慢性疼痛患者的认知障碍。但是,与针灸对疼痛和认知障碍的共同治愈作用相关的详细机制尚未揭示。在本研究中,我们发现在GB30和GB34穴位进行针刺治疗可改善与PSNL诱发的神经性疼痛模型相关的伤害性行为和认知障碍。使用机器学习的决策树算法进行分析时,针灸组被分类为与PSNL组不同。我们还发现,针刺可以通过增加LTP以及海马中NR2B和GluR1的表达水平来增强突触可塑性,而PSNL组中受损。另外,通过针刺处理可增强CaMKII和突触蛋白如pPSD-95和pSyn-1的表达,突触可塑性的指标。
,开发周围神经损伤中枢敏被认为有助于慢性疼痛和在临床前模型和患者的认知功能的不利影响 。在这里,我们观察到具有长时间机械异常性疼痛的PSNL小鼠在进行Y迷宫和NOR测试时,其工作和识别记忆受到损害,与最近的研究一致。相反,针灸治疗(AP1)可以改善记忆功能以及机械性异常性疼痛。其他穴位或非穴位都没有导致这种改善,这表明GB30和GB34穴位对这些治疗效果具有特异性。接下来,使用机器学习中的随机决策森林分类器来确定是否可以基于疼痛和认知行为来预测针灸的治疗效果。机器学习分类器是机器学习技术的特定应用程序,它使用各个功能来预测给定示例所属的预先分配的类。给定模型的成功可通过检查其忠实地预测组成员身份的程度来评估,还可以比较功能集之间的性能。我们发现,针刺治疗的小鼠不仅被归类为与PSNL小鼠不同的组,而且还向假手术小鼠转移。相反,在与PSNL组聚集的对照点进行针灸治疗的小鼠。
海马在认知功能中起关键作用。在患有认知障碍的阿尔茨海默氏病啮齿动物模型中发现海马体积减少和突触可塑性改变。在慢性疼痛中也观察到了这种海马体的变化,突触可塑性的降低可能与疼痛并发的认知障碍有关。LTP是一个与活动有关的突触强度,基础信息存储和记忆创建方面的变化的细胞模型。最近,在几种疼痛模型中报告了海马LTP降低,例如周围神经损伤引起的认知障碍小鼠。与以前的研究一致,我们还发现,在患有认知障碍的PSNL小鼠的海马CA1中测量LTP会降低。此外,我们发现,与PSNL诱导的神经性疼痛小鼠相比,针灸增强了LTP。最近的慢性疼痛实验报告说,针刺可调节突触功能并调节认知障碍,但尚未提出分子机制。因此,我们研究了与针刺治疗有关的认知功能和疼痛改善相关的潜在机制,重点研究了海马体中与LTP相关的分子机制。
在Schaffer侧支CA1突触中LTP的诱导和维持是NMDA和AMPA受体依赖性的。NR2B通过增加钙增加的CaMKII活性2 +流入突触,然后增加在突触的GluR1的表达,因此介导许多重要的脑功能,包括认知,学习和记忆。海马神经元中NR2B,GluR1,CaMKII和突触蛋白的下调与阿尔茨海默氏病(AD)动物模型中的学习记忆缺陷和认知功能障碍有关。如最近的一项AD研究所示,在PSNL诱导的神经性疼痛模型中,海马NR2B的表达降低。报道了神经病性疼痛小鼠海马CA3区的GluR1以及共表达脑源性神经营养因子和GluR1的神经元表达下调,这也与疼痛相关的合并症有关。与这些研究一致,我们的结果表明,PSNL神经性疼痛模型的CA1和CA3中海马NR2B和GluR1明显下调,而针灸治疗可以挽救它们。有趣的是,相关分析表明,NR2B和GluR1的表达水平与认知功能呈正相关,而与疼痛行为呈负相关。这表明,AP1组中NR2B和GluR1的增加可对疼痛和合并症认知功能障碍的恢复起关键作用。此外,我们表明,针刺可增强海马中pCaMKII和tCaMKII的表达水平,而在神经性疼痛模型中则降低。CaMKII的激活对于海马中LTP的诱导至关重要,并且它与活性依赖性突触的增强有关。PKC-γ通过增加的细胞内Ca 2+内流而被激活,但是针灸治疗并没有改善。Fernándezde Sevilla和Buño等。提示CaMKII在启动乙酰胆碱介导的NMDAR和AMPAR介导的传递的增强中起重要作用,但PKC在启动乙酰胆碱介导的仅NMDAR介导的传递的增强中可能是必不可少的。因此,我们推测海马中CaMKII和PKC表达模式的差异可能与针灸治疗的胆碱能机制有关,需要进一步研究。
最后,我们证明了PSNL模型小鼠海马中突触蛋白(例如Syn-1和PSD-95)的显着改变。Syn-1是突触前的终末特异性标记物,参与囊泡的运输,释放和再循环,并被认为在钙依赖性神经递质释放过程中发挥重要作用,例如谷氨酸 。PSD-95是突触后的结构蛋白,并通过CaMKII的激活而增加,而CaMKII在突触可塑性中起着至关重要的作用。在这项研究中,我们发现针灸治疗可以恢复PSNL诱导的神经性疼痛海马中磷酸化的Syn-1和PSD-95的水平降低。因此,我们的结果表明,针刺可以通过增加谷氨酸受体和突触蛋白的表达来调节谷氨酸的传递,从而改善伤害感受和认知障碍。
众所周知,神经元之间的突触是神经连接和神经可塑性的结构基础。在最近的研究中,在神经性疼痛小鼠中,CA1锥体神经元的总树突长度,树突分支的数量和基础和顶端树突的脊柱密度显着降低了。因此,需要进一步的研究来探索针灸是否可以改变与突触可塑性有关的CA1锥体神经元的形态。
这项研究的局限性是未知,周围神经系统的变化如何影响大脑。针灸通过增加肌肉中腺苷和单磷酸腺苷的释放显示出镇痛作用,而这种作用在腺苷A1受体敲除小鼠中消失了。针刺镇痛中也介导了局部激活的ERK信号通路。但是,由于这项研究的重点是中枢神经系统,因此我们没有密切观察针灸治疗后周围神经系统中腺苷,ATP或ERK途径的变化。另外,已知在神经元以及至少部分神经胶质细胞或小神经胶质细胞中,对代谢型A1受体的刺激与脊髓和上脊髓水平的慢性疼痛的调节有关。此外,在腺苷A1受体敲除小鼠中海马CA1区的LTP被抑制。需要进一步的研究来定义局部组织中的腺苷A1受体与海马中的谷氨酸受体之间的关系。
- Happy Halloween -
小结
总之,我们证明了针灸治疗可以同时改善异常性疼痛和合并症认知障碍,并且针灸的治疗效果与海马NR2B和GluR1表达的增加有关。针刺还增强了该区域的pCaMKII和tCaMKII以及突触前和突触后蛋白。最后,这些分子变化有助于海马CA1中LTP的增加。这些结果可能暗示,针灸可能是控制疼痛和合并症的潜在障碍之一,同时还需要进一步的临床试验来证明。
文献来源
Acupuncture Improves Comorbid Cognitive Impairments Induced by Neuropathic Pain in Mice. Front Neurosci, 2019. 13: p. 995.







 中国中医药
中国中医药


